La aparición de las primeras formas de vida sobre la Tierra es uno de los problemas que más ha interesado a los biólogos desde hace siglos y, con toda seguridad, seguirá siendo un problema abierto aún durante mucho tiempo. La falta de registros fósiles de épocas tan antiguas hace que el estudio de este proceso tenga que basarse exclusivamente en evidencias indirectas y extrapolaciones, que dejan mucho margen de error posible y permiten múltiples interpretaciones.
Aunque siguen siendo muchos los hechos que se desconocen, se empiezan a tener algunas ideas relativamente claras. Por una parte, algunas evidencias geológicas han retrasado la aparición de la vida hasta un momento más antiguo de lo que se creía hace algunas décadas, hasta hace unos 3.900 millones de años de modo que el periodo de evolución prebiótica, más largo, resulta más verosimil. También se ha mejorado mucho en el conocimiento de las condiciones geológicas y ambientales de nuestro planeta, y de la geoquímica de cometas y asteroides, que pudieron tener un papel importante en la aparición de las primeras moléculas orgánicas.
Algunos aspectos de la evolución prebiótica van siendo establecidos poco a poco, mientras que otros siguen siendo objeto de profundas controversias. Así las cosas, el panorama actual está configurado por una amplia gama de hipótesis, en su mayor parte apoyadas por una evidencia relativamente débil.
En cualquier caso, es ampliamente aceptado que la evolución prebiótica tuvo que cumplir ciertos requisitos:
En cualquier caso, es ampliamente aceptado que la evolución prebiótica tuvo que cumplir ciertos requisitos:
- Se trata de un proceso evolutivo, regido por los principios de la Selección Natural.
- Necesariamente, esta evolución tuvo que atravesar ciertos pasos, cuyos resultados observamos en los organismos actuales:
- Formación de las moléculas orgánicas.
- Adquisición, por parte de algunas de esas moléculas, de funciones características de la vida: capacidad catalítica, capacidad para almacenar y transmitir información.
- Asociación coordinada de esas moléculas y de las funciones realizadas por ellas en sistemas "discretos", es decir, separados de su entorno.
- Aparición de las primeras protocélulas.
La Tierra primitiva
En la actualidad conviven dos grandes tendencias teóricas relacionadas con el posible origen de la vida en nuestro planeta: las que consideran que todo el proceso tuvo lugar en la Tierra y las que opinan que cuerpos exteriores (meteoritos o cometas) pudieron jugar un papel significativo en la formación o evolución, al menos, de las sustancias químicas necesarias para la aparición de la vida. En cualquier caso, para ambos grupos de teorías sigue siendo fundamental conocer las condiciones ambientales que se daban en nuestro planeta cuando empezaron a sintetizarse los primeros compuestos orgánicos.
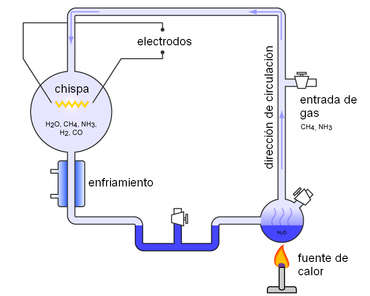
Los modelos originales acerca de la síntesis abiótica de sustancias orgánicas (el clásico experimento de Miller y Urey que consiguió sintetizar aminoácidos en presencia de una fuente de energía a partir de sustancias inorgánicas) suponían que la composición de la atmósfera primitiva de la Tierra debió ser muy reductora. Con los datos que se poseen en la actualidad, parece bastante improbable que esto fuera realmente así. Más bien se considera que el vapor de agua debió ser el componente mayoritario de esa atmósfera primitiva, junto a una considerable cantidad de dióxido de carbono. Sin embargo, sí que parece posible que existiera también una cierta cantidad de metano (entre 10 y 100 ppm). La presencia de este gas es particularmente importante, porque sus reacciones desencadenadas por la luz pueden dar lugar a la formación de HCN y H2CO, compuestos que pueden intervenir en la síntesis de aminoácidos, glúcidos y ácidos nucleicos. Esta composición debió ir variando paulatinamente durante algunos cientos de millones de años: la descomposición del metano dio lugar a hidrógeno, que escaparía de la atmósfera, y por reacción con otras sustancias, a CO2. Este gas, por su parte, iría desapareciendo de la atmósfera a medida que se incorporaba a la composición de las rocas carbonatadas. En cuanto a la presencia de oxígeno, este gas se forma en ciertas reacciones que pudieron tener lugar en una atmósfera como la descrita, pero no pudo acumularse en la atmósfera debido a que reaccionaría con las sustancias reducidas emitidas por los volcanes.
Por otra parte, el clima en la era Arcaica (periodo geológico durante el cual se produjo la evolución prebiótica) fue también un factor fundamental para la aparición de la vida. Durante esta época la temperatura tuvo que ser, forzosamente, suficientemente elevada para permitir las reacciones de síntesis orgánicas. Esta necesidad se enfrenta al llamado "problema del Sol joven frío": por aquel entonces nuestro Sol emitía un 30% menos de energía que en la actualidad, lo que habría provocado una temperatura excesivamente baja, congelando todo el planeta e impidiendo la evolución química. Posiblemente, la temperatura pudo mantenerse dentro de los márgenes apropiados para la vida gracias al efecto invernadero producido, en primer lugar, por los gases presentes en la atmósfera original (CH4, CO2) y, a medida que éstos se iban consumiendo, por la acción de los organismos vivos, en particular de los metanógenos.
La aparición de las primeras moléculas orgánicas
Uno de los principales puntos de discusión en el ámbito del origen de la vida es la formación de las primeras sustancias orgánicas. Existen dos grandes tendencias: la de quienes suponen que tales moléculas se originaron en nuestro planeta, posiblemente en entornos muy concretos caracterizados por temperaturas elevadas, presencia de agua y minerales y energía química abundante, y la de quienes piensan que esas condiciones son muy improbables, y que tales sustancias pudieron llegar a la Tierra como consecuencia del choque de cometas o meteoritos. Hay evidencias indirectas a favor de ambas hipótesis: las surgencias oceánicas, por ejemplo, son buenos candidatos para la formación de compuestos orgánicos, y se han encontrado sustancias de esta naturaleza (aminoácidos, glúcidos, nucleótidos...) en cometas y meteoritos. Por otra parte, nada indica que las dos ideas sean incompatibles entre sí, lo que ha favorecido que últimamente se vaya imponiendo una teoría "sincrética".
Por otra parte, el clima en la era Arcaica (periodo geológico durante el cual se produjo la evolución prebiótica) fue también un factor fundamental para la aparición de la vida. Durante esta época la temperatura tuvo que ser, forzosamente, suficientemente elevada para permitir las reacciones de síntesis orgánicas. Esta necesidad se enfrenta al llamado "problema del Sol joven frío": por aquel entonces nuestro Sol emitía un 30% menos de energía que en la actualidad, lo que habría provocado una temperatura excesivamente baja, congelando todo el planeta e impidiendo la evolución química. Posiblemente, la temperatura pudo mantenerse dentro de los márgenes apropiados para la vida gracias al efecto invernadero producido, en primer lugar, por los gases presentes en la atmósfera original (CH4, CO2) y, a medida que éstos se iban consumiendo, por la acción de los organismos vivos, en particular de los metanógenos.
La aparición de las primeras moléculas orgánicas
Uno de los principales puntos de discusión en el ámbito del origen de la vida es la formación de las primeras sustancias orgánicas. Existen dos grandes tendencias: la de quienes suponen que tales moléculas se originaron en nuestro planeta, posiblemente en entornos muy concretos caracterizados por temperaturas elevadas, presencia de agua y minerales y energía química abundante, y la de quienes piensan que esas condiciones son muy improbables, y que tales sustancias pudieron llegar a la Tierra como consecuencia del choque de cometas o meteoritos. Hay evidencias indirectas a favor de ambas hipótesis: las surgencias oceánicas, por ejemplo, son buenos candidatos para la formación de compuestos orgánicos, y se han encontrado sustancias de esta naturaleza (aminoácidos, glúcidos, nucleótidos...) en cometas y meteoritos. Por otra parte, nada indica que las dos ideas sean incompatibles entre sí, lo que ha favorecido que últimamente se vaya imponiendo una teoría "sincrética".
De acuerdo con esta idea, la vida podría haber surgido en un tipo de ambientes muy concreto, denominado "FAIR" (Fire And Ice Reactors). Un entorno FAIR estaría constituido por una zona de lagunas intermareales, situada en una región subpolar, suficientemente cálida como para no estar permanentemente congelada, pero suficientemente fría como para permitir, periódicamente, el contacto entre agua y hielo. En esa zona debería producirse mezcla entre agua dulce, por ejemplo procedente de la lluvia, y agua marina, para dar lugar a un gradiente iónico que permitiera la solubilización de los fosfatos y ciertos tipos de reacciones químicas.

Resultaría necesaria también la presencia de fumarolas y aguas termales, que proporcionarían un flujo abundante de minerales, entre ellos arcillas. Por último, la caída en la zona de meteoritos o cometas podría representar otra fuente de materia orgánica.
Podría parecer que se trata de condiciones tan restrictivas que hacen muy improbable la existencia de tales zonas. Sin embargo, incluso en la actualidad pueden encontrarse ámbitos como estos FAIR en zonas como la península de Kamchatka, Alaska o Islandia, y cabe suponer que en el pasado, sin la competencia colonizadora de otros organismos, pudieron resultar mucho más frecuentes.
Los procesos de síntesis de biomoléculas debieron dar lugar a varios tipos de sustancias que luego serían necesarias para dar lugar a los primeros seres vivos:
Otra de las cuestiones importantes en esta fase fue la fuente de energía que permitió, en primer lugar, la propia síntesis de moléculas orgánicas y más adelante que los sistemas prebióticos pudieran llevar a cabo reacciones endotérmicas necesarias para desarrollar su metabolismo. Aunque hay varios candidatos posibles para actuar como fuentes de energía, todas ellos presentan puntos débiles que los hacen improbables por sí mismos como sistema de suministro energético para esta etapa de la evolución, por lo que se ha sugerido que los primeros sistemas prebióticos pudieron utilizar una combinación de varias de estas fuentes energéticas disponibles a su alrededor:
En una primera fase, moléculas de polifosfato como las que se producen en las aguas termales pudieron proporcionar energía para la síntesis de nucleótidos trifosfato que formaran el ARN o alguna molécula relacionada. Los experimentos que recrean las condiciones arcaicas parecen demostrar que la adenina es la base nitrogenada que más fácilmente se forma, lo que explicaría que el ATP sea la molécula más utilizada por todos los organismos como moneda energética. En esta fase pudo producirse un acoplamiento entre reacciones metabólicas exotérmicas que implicaran ácidos orgánicos y aminoácidos, compuestos seguramente abundantes y polímeros de nucleótidos que serían los primeros sistemas replicadores, por lo que la cantidad de cadenas de nucleótidos pudo crecer bastante rápidamente.
El paso de la "sopa de moléculas" a los siguientes niveles de organización prebióticos pudo resultar bastante complicado. Antes de poder ser considerado célula, un sistema biológico primitivo tuvo que reunir al menos tres características complejas incluso por separado: presencia de membrana, capacidad de replicación y metabolismo. Las membranas permiten que los "protoorganismos" actúen como unidades estructurales separadas de su entorno, la capacidad de replicación proporciona la posibilidad de conservar y transmitir la información necesaria para organizar estructural y funcionalmente el organismo y el metabolismo hace posible dicho funcionamiento, proporcionando los elementos constituyentes y la energía necesaria.
De las tres características, la membrana es la más fácil de adquirir: las membranas lipídicas pueden formarse espontáneamente a partir de moléculas fáciles de sintetizar, o procedentes de meteoritos. Además, es posible imaginar otros mecanismos para mantener asociadas las moléculas necesarias para la replicación y el metabolismo. Por ejemplo, se ha propuesto que las arcillas y otros sistemas minerales pudieron jugar ese papel en fases tempranas de la evolución prebiótica.
En cambio, resulta muy difícil imaginar que la capacidad de replicación y la actividad metabólica pudieran evolucionar por separado: los elementos de una ruta metabólica no podrían haberse autoorganizado, ni tendrían capacidad para evolucionar, mientras que la formación de un conjunto de replicadores funcional, sin ninguna ventaja metabólica, resulta altamente improbable. De este modo, lo que parece más probable es que ambas capacidades evolucionaran simultáneamente, de forma que el proceso consistiera en la selección metabólica de un conjunto de replicadores, posiblemente formados por ARN o por alguna sustancia similar.

Resultaría necesaria también la presencia de fumarolas y aguas termales, que proporcionarían un flujo abundante de minerales, entre ellos arcillas. Por último, la caída en la zona de meteoritos o cometas podría representar otra fuente de materia orgánica.
Podría parecer que se trata de condiciones tan restrictivas que hacen muy improbable la existencia de tales zonas. Sin embargo, incluso en la actualidad pueden encontrarse ámbitos como estos FAIR en zonas como la península de Kamchatka, Alaska o Islandia, y cabe suponer que en el pasado, sin la competencia colonizadora de otros organismos, pudieron resultar mucho más frecuentes.
Los procesos de síntesis de biomoléculas debieron dar lugar a varios tipos de sustancias que luego serían necesarias para dar lugar a los primeros seres vivos:
- Aminoácidos: diferentes estudios demuestran que en las condiciones ambientales descritas se pudieron llegar a formar hasta diez de los aminoácidos presentes en las proteínas. Curiosamente, esos mismos aminoácidos se han podido encontrar en la composición de los meteoritos y son, además, los que forman parte del código genético primitivo que aún puede rastrearse en los organismos actuales. Este fenómeno, denominado triple convergencia, sugiere que los primeros organismos pudieron utilizar solo esos aminoácidos en sus proteínas.
- Nucleótidos: la síntesis abiótica de las bases nitrogenadas es posible en las condiciones ambientales que se supone que existían en la era Arcaica, pero la formación de nucleótidos parece ser un proceso bastante más difícil, por lo que se ha sugerido que las primeras moléculas informativas no fueron los ácidos nucleico, sino algunos compuestos parecidos a ellos a partir de los cuales pudieron evolucionar.
- Lípidos: los lípidos anfipáticos son elementos fundamentales de las membranas y, por lo tanto, esenciales para la evolución de las células. Los restos extraídos de un famoso meteorito, el de Murchison, contenían lípidos anfipáticos que, al ser mezclados con agua, formaron espontáneamente vesículas lipídicas. También se ha podido demostrar la posibilidad de que lípidos anfipáticos de este tipo se produzcan en las surgencias oceánicas.
Otra de las cuestiones importantes en esta fase fue la fuente de energía que permitió, en primer lugar, la propia síntesis de moléculas orgánicas y más adelante que los sistemas prebióticos pudieran llevar a cabo reacciones endotérmicas necesarias para desarrollar su metabolismo. Aunque hay varios candidatos posibles para actuar como fuentes de energía, todas ellos presentan puntos débiles que los hacen improbables por sí mismos como sistema de suministro energético para esta etapa de la evolución, por lo que se ha sugerido que los primeros sistemas prebióticos pudieron utilizar una combinación de varias de estas fuentes energéticas disponibles a su alrededor:
En una primera fase, moléculas de polifosfato como las que se producen en las aguas termales pudieron proporcionar energía para la síntesis de nucleótidos trifosfato que formaran el ARN o alguna molécula relacionada. Los experimentos que recrean las condiciones arcaicas parecen demostrar que la adenina es la base nitrogenada que más fácilmente se forma, lo que explicaría que el ATP sea la molécula más utilizada por todos los organismos como moneda energética. En esta fase pudo producirse un acoplamiento entre reacciones metabólicas exotérmicas que implicaran ácidos orgánicos y aminoácidos, compuestos seguramente abundantes y polímeros de nucleótidos que serían los primeros sistemas replicadores, por lo que la cantidad de cadenas de nucleótidos pudo crecer bastante rápidamente.
El paso de la "sopa de moléculas" a los siguientes niveles de organización prebióticos pudo resultar bastante complicado. Antes de poder ser considerado célula, un sistema biológico primitivo tuvo que reunir al menos tres características complejas incluso por separado: presencia de membrana, capacidad de replicación y metabolismo. Las membranas permiten que los "protoorganismos" actúen como unidades estructurales separadas de su entorno, la capacidad de replicación proporciona la posibilidad de conservar y transmitir la información necesaria para organizar estructural y funcionalmente el organismo y el metabolismo hace posible dicho funcionamiento, proporcionando los elementos constituyentes y la energía necesaria.
De las tres características, la membrana es la más fácil de adquirir: las membranas lipídicas pueden formarse espontáneamente a partir de moléculas fáciles de sintetizar, o procedentes de meteoritos. Además, es posible imaginar otros mecanismos para mantener asociadas las moléculas necesarias para la replicación y el metabolismo. Por ejemplo, se ha propuesto que las arcillas y otros sistemas minerales pudieron jugar ese papel en fases tempranas de la evolución prebiótica.
En cambio, resulta muy difícil imaginar que la capacidad de replicación y la actividad metabólica pudieran evolucionar por separado: los elementos de una ruta metabólica no podrían haberse autoorganizado, ni tendrían capacidad para evolucionar, mientras que la formación de un conjunto de replicadores funcional, sin ninguna ventaja metabólica, resulta altamente improbable. De este modo, lo que parece más probable es que ambas capacidades evolucionaran simultáneamente, de forma que el proceso consistiera en la selección metabólica de un conjunto de replicadores, posiblemente formados por ARN o por alguna sustancia similar.
El mundo de ARN
Los sistemas biológicos actuales funcionan gracias a la interdependencia de ácidos nucleicos y proteínas: los ácidos nucleicos almacenan y transmiten la información necesaria para sintetizar las proteínas, mientras que las proteínas son necesarias para la replicación de los ácidos nucleicos. Parece evidente que los primeros sistemas biológicos tuvieron que ser más sencillos, de modo que inicialmente debieron contar solo con uno de estos dos tipos de moléculas. El problema de cuál de los dos tipos de moléculas apareció antes no se resolvió definitivamente hasta que se descubrió que algunos ARN tenían, además, capacidad catalítica (ribozimas).
El proceso hipotético que dio lugar a un mundo de ARN pudo ser el siguiente: en primer lugar, debió tener lugar la formación de nucleótidos a partir de precursores más pequeños. La unión de los nucleótidos para formar cadenas debió tener lugar en presencia de catalizadores, pero dado que por esa época no existían las proteínas, es posible que ese papel lo desempeñaran iones metálicos.
Las cadenas de nucleótidos con capacidad catalítica son de tamaño considerable. Su formación pudo tener lugar por adición de nucleótidos individuales o por unión de cadenas más cortas, pero finalmente algunas de esas cadenas debieron adquirir la capacidad para replicarse a sí mismas sin necesidad de intervención enzimática. Esta capacidad permitió tanto incrementar la capacidad catalítica de los ARN como crear una "sopa" de ácidos nucleicos capaz de evolucionar mediante errores en el proceso de copia y selección natural, lo que condujo al "mundo de ARN", en el que se mezclaban moléculas de ARN capaces de catalizar su propia replicación con otras que presentaban otras capacidades catalíticas.
Un mundo de ARN exige que este tipo de molécula sea capaz de realizar una amplia gama de funciones bioquímicas, más restringidas que las que actualmente realizan las proteínas, pero en todo caso mucho más extendida que las que ahora realizan las escasas ribozimas que se conocen. Se han encontrado evidencias indirectas que pueden apoyar la capacidad de los ARN para actuar como enzimas, como la abundancia de cofactores enzimáticos basados en nucleótidos (que sugeriría que las enzimas "conservaron" la capacidad catalítica de estas moléculas de sus precursores, las ribozimas); también se han propuesto mecanismos de reacción hipotéticos que podrían explicar cómo un ARN podría catalizar su replicación, sin necesidad de proteínas.
El mayor problema para explicar un mundo de ARN, sin embargo, sigue siendo resolver el "acoplamiento" entre los ARN y las proteínas. Para resolverlo se ha propuesto un mecanismo que tiene en cuenta la facilidad con la que se producen aminoácidos en procesos de síntesis abiótica. Las ribozimas podrían haber dispuesto, a su alrededor, de una buena cantidad de aminoácidos que inicialmente pudieron utilizar como cofactores. La unión de aminoácidos en cadenas pudo permitir mejorar la precisión y la complejidad de los procesos de catálisis, por lo que habría sido seleccionado positivamente, evolucionando hacia un mundo de ácidos nucleicos y proteínas.
Los sistemas biológicos actuales funcionan gracias a la interdependencia de ácidos nucleicos y proteínas: los ácidos nucleicos almacenan y transmiten la información necesaria para sintetizar las proteínas, mientras que las proteínas son necesarias para la replicación de los ácidos nucleicos. Parece evidente que los primeros sistemas biológicos tuvieron que ser más sencillos, de modo que inicialmente debieron contar solo con uno de estos dos tipos de moléculas. El problema de cuál de los dos tipos de moléculas apareció antes no se resolvió definitivamente hasta que se descubrió que algunos ARN tenían, además, capacidad catalítica (ribozimas).
El proceso hipotético que dio lugar a un mundo de ARN pudo ser el siguiente: en primer lugar, debió tener lugar la formación de nucleótidos a partir de precursores más pequeños. La unión de los nucleótidos para formar cadenas debió tener lugar en presencia de catalizadores, pero dado que por esa época no existían las proteínas, es posible que ese papel lo desempeñaran iones metálicos.
Las cadenas de nucleótidos con capacidad catalítica son de tamaño considerable. Su formación pudo tener lugar por adición de nucleótidos individuales o por unión de cadenas más cortas, pero finalmente algunas de esas cadenas debieron adquirir la capacidad para replicarse a sí mismas sin necesidad de intervención enzimática. Esta capacidad permitió tanto incrementar la capacidad catalítica de los ARN como crear una "sopa" de ácidos nucleicos capaz de evolucionar mediante errores en el proceso de copia y selección natural, lo que condujo al "mundo de ARN", en el que se mezclaban moléculas de ARN capaces de catalizar su propia replicación con otras que presentaban otras capacidades catalíticas.
 |
| Cofactores enzimáticos derivados de nucleótidos |
El mayor problema para explicar un mundo de ARN, sin embargo, sigue siendo resolver el "acoplamiento" entre los ARN y las proteínas. Para resolverlo se ha propuesto un mecanismo que tiene en cuenta la facilidad con la que se producen aminoácidos en procesos de síntesis abiótica. Las ribozimas podrían haber dispuesto, a su alrededor, de una buena cantidad de aminoácidos que inicialmente pudieron utilizar como cofactores. La unión de aminoácidos en cadenas pudo permitir mejorar la precisión y la complejidad de los procesos de catálisis, por lo que habría sido seleccionado positivamente, evolucionando hacia un mundo de ácidos nucleicos y proteínas.

