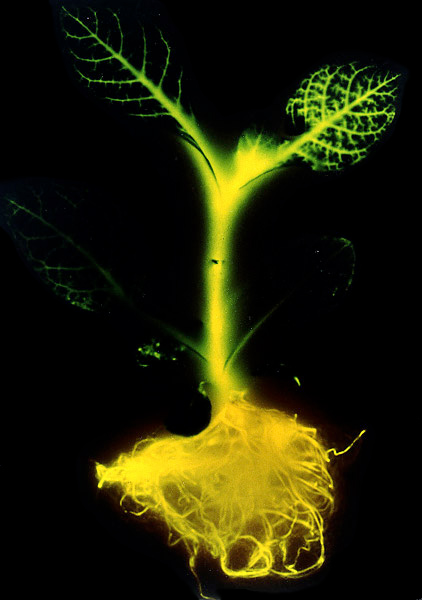
La alteración de las características genéticas de los organismos es una actividad que el ser humano ha practicado desde la antigüedad. Tradicionalmente, la búsqueda de características biológicas útiles para el hombre se ha hecho utilizando mecanismos que no implicaban la alteración directa del material genético. Sin embargo, actividades tales como la selección de los organismos que presentan características interesantes, el cruzamiento de individuos para obtener otros con una combinación más apropiada, o la hibridación de variedades son, en realidad, técnicas que inciden directamente sobre el genoma de esos organismos.
En el siglo XX, el descubrimiento de la naturaleza química del material genético abrió la puerta a la utilización de nuevas técnicas. Primero fue la mutagénesis dirigida, consistente en tratar a los organismos con un agente mutágeno y seleccionar aquellos que, como resultado de las mutaciones, presentan características interesantes. Más adelante, a partir de los años 70' del pasado siglo, los conocimientos aportados por la Biología Molecular permitieron la manipulación directa del material genético.
Habitualmente se suelen confundir los conceptos de manipulación genética y de ingeniería genética. En realidad, la manipulación genética "indirecta" ha existido siempre, como lo muestra la gran diferencia que existe entre los organismos "domesticados" (ya sean animales o plantas) y sus ancestros "silvestres". Los resultados de la selección artificial en Brassica se muestran en la siguiente imagen.
La ingeniería genética, por su parte, es el término que suele utilizarse para la alteración directa y dirigida del material genético de un ser vivo.
La naturaleza química del ADN, y la presencia en los organismos de una gran diversidad de enzimas que son capaces de actuar sobre esta molécula, han permitido a los investigadores utilizar estas máquinas moleculares para alterar de un modo controlado la información genética de los organismos. Gracias a sus propiedades, los biotecnólogos son capaces de manipular el ADN de varios modos diferentes:
- Extracción: consiste en separar el material genético de una célula del resto de los componentes de la misma. La extracción es el paso inicial, imprescindible, para cualquier otro proceso de manipulación.
- Corte del ADN: las moléculas de ADN tienen un tamaño enorme, y contienen grandes cantidades de información. Para poder manipularlas de forma más o menos eficaz es necesario trocearlas, para poder alterar solo aquellos fragmentos que tienen los genes que nos interesan. El corte de las moléculas de ADN se realiza utilizando enzimas extraidas de algunos organismos, que actúan, en algunos casos, cortando la molécula en fragmentos de cierto tamaño, y en otros cortando el ADN en zonas concretas, en las que se presentan ciertas secuencias reconocidas por las enzimas de corte (restrictasas).
- Empalme del ADN: los fragmentos de ADN que han sido obtenidos en los procesos anteriores pueden unirse entre sí o con otros, con ayuda de la ligasa, una de las enzimas que participan en el proceso de replicación de ADN. Esto permite unir un fragmento que se haya seleccionado a otra molécula de ADN, incorporándole las características de ese fragmento.
- Transformación: reintroducción del material genético manipulado en un organismo para que lo exprese y de lugar a la proteína que se buscaba conseguir.
La ingeniería genética incluye una gran variedad de técnicas y actividades, aunque como ejemplos se pueden citar algunas de gran interés:
- Clonación génica: consiste en la identificación, extracción y aislamiento de un gen para integrarlo en el genoma de un organismo diferente. De este modo se consigue que un organismo, normalmente fácil de cultivar en procesos industriales, sea capaz de sintetizar compuestos propios de otros. Un ejemplo ya tradicional de este tipo de técnicas es la cloanción del gen de la insulina: se consiguió aislar el gen humano que codifica para la producción de la insulina, e introducirlo en Escherichia coli, un organismo muy fácil de cultivar, de modo que la bacteria produce, en instalaciones industriales y en grandes cantidades, insulina que puede ser utilizada como medicamento en el tratamiento de la diabetes.


- Pruebas de ADN: se trata de identificar la procedencia de fragmentos de ADN de origen desconocido, relacionándolos con una muestra cuyo origen sí se conoce. Las pruebas de ADN se basan en el principio de complementariedad de bases, ya que consisten, en definitiva, de asociar una cadena de la molécula problema (la que queremos investigar) con otra que suponemos que es su complementaria. Para poder realizar este tipo de ensayos es necesario contar con una cantidad relativamente grande de ADN problema, lo que se consigue gracias a la técnica llamada "PCR" (reacción en cadena de la polimerasa), que permite multiplicar fácilmente pequeñas cantidades iniciales de ADN. Las aplicaciones prácticas de las pruebas de ADN son muy variadas, y van desde la identificación de personas (por ejemplo, en los casos de investigación forense) hasta el estudio evolutivo, por comparación de las secuencias de ADN de especies extinguidas y actuales, para determinar el grado de parentesco, pasando por aplicaciones médicas como la identificación de genes responsables de la aparición de ciertas enfermedades.
- Clonación de individuos completos: en este caso se pretende conseguir un organismo completo idéntico en todos los aspectos determinados genéticamente a uno que ya existía. En vegetales es un procedimiento extraordinariamente sencillo, hasta el punto de que cualquier aficionado a la jardinería lo lleva a cabo cuando reproduce una planta mediante esquejes. En animales el proceso es bastante complejo, especialmente en cordados, porque las células del organismo adulto están "diferenciadas", es decir, han bloqueado la expresión de algunos de sus genes. La primera clonación de vertebrados se llevó a cabo en la década de los 60' del siglo XX, en un tipo de sapo africano. Sin embargo, no hay duda de que el caso de clonación más conocido es el de la oveja Dolly, que fue el primer mamífero con el que tuvo éxito esta técnica.
- Terapia génica: consiste en insertar un gen funcional en las células de un paciente humano para corregir un defecto genético o para dotar a las células de una nueva función.
Clonación de un gen
La clonación génica consiste en extraer un gen de un organismo e introducirlo en otro distinto, de modo que el segundo sea capaz de producir la proteína codificada por ese gen. Este tipo de técnicas se utilizan para producir medicamentos u otras proteínas de interés (por ejemplo anticuerpos), pero también para que el organismo genéticamente modificado adquiera características de las que inicialmente carecía. En este sentido se han utilizado técnicas de ingeniería genética, por ejemplo, para conseguir que algunas frutas u hortalizas maduren más lentamente, con lo que su duración es mayor.
La clonación génica pasa por las siguientes etapas:
- Extracción del ADN del organismo cuyo gen se quiere clonar.
- Fragmentación del ADN extraído.
- Identificación/aislamiento del gen buscado.
- Introducción del gen en el organismo que va a producir la proteína clonada.
- Identificación y selección de los organismos transformados que poseen el gen clonado.
La extracción del ADN es un proceso relativamente sencillo, gracias a que el ADN es una molécula bastante resistente a los tratamientos químicos. El procedimiento incluye la rotura (lisis) de las células, seguida de una centrifugación en la que se eliminan los restos celulares de mayor tamaño. La separación del ADN del resto de los compuestos solubles se realiza por precipitación con alcohol isopropílico. Una nueva centrifugación da lugar a un precipitado de ADN prácticamente puro.
El segundo paso de la clonación es la individualización del gen que interesa clonar. El material genético de partida es el genoma completo de un organismo, millones de bases de las que interesan apenas unas mil. Además, no existe un modo directo de encontrar un gen entre tanto material genético. En todo caso, lo primero que hay que hacer con ese material genético es "trocearlo", utilizando para ello enzimas capaces de cortar la molécula de ADN, que reciben el nombre de restrictasas.
Existen varios tipos de restrictasas. Algunas de ellas son "inespecíficas", lo que significa que cortan la cadena de ADN independientemente de la secuencia que éste presente. En ese caso el corte puede producirse al azar o cada cierto número de nucleótidos. Sin embargo, las restrictasas más interesantes para la ingeniería genética son las que reconocen ciertas secuencias de ADN y cortan la molécula precisamente por esas zonas.
Las secuencias reconocidas por las restrictasas suelen ser palindrómicas, es decir, capicúas, y en muchos casos las enzimas las cortan dejando cadenas con extremos de diferente longitud, como se aprecia en la imagen:
Los extremos de este corte reciben el nombre de "cohesivos" porque actúan, en cierto sentido, como "tiras de velcro": las zonas no apareadas se complementan con las de otra molécula de ADN cortadas con la misma enzima, de modo que si se ponen juntos dos fragmentos de ADN cortados con la misma restrictasa (y esta deja fragmentos cohesivos), los extremos de dichos fragmentos se unirán entre sí espontáneamente, permitiendo que vuelvan a unirse.
En general, el ADN aislado se corta con una enzima de restricción que proporcione extremos cohesivos. Si se conoce la secuencia del gen, se trata de seleccionar una restrictasa que no corte la secuencia del gen. Existe un número considerable de restrictasas, cuya secuencia de corte es conocida, lo que facilita bastante el trabajo en este caso.
El gen que se busca es uno de los miles de fragmentos que se han obtenido en el proceso de corte. La individualización de los fragmentos de ADN se hace utilizando técnicas de separación, en especial la electroforesis en gel de agarosa. Esta técnica aprovecha el hecho de que el ADN esté cargado eléctricamente a pH fisiológico. Si se aplica un campo eléctrico a una mezcla de moléculas de ADN éstas se mueven hacia el polo positivo, a una velocidad que es inversamente proporcional a su tamaño. De este modo, haciendo correr esas moléculas a través de un gel durante un tiempo fijo es posible separar las moléculas en función de su longitud.
La electroforesis en gel permite separar entre sí los fragmentos de ADN según su tamaño, lo que facilita su identificación. Sin embargo, no es suficiente; aún es posible que haya cientos de fragmentos de ADN de tamaño similar al que se busca, por lo que sigue siendo necesario localizar el gen concreto que interesa en el proceso. Básicamente, se utilizan dos técnicas para conseguirlo. La más antigua es la elaboración de una "genoteca", aunque el desarrollo de técnicas más modernas ha permitido utilizar "sondas" para identificar genes.
La estrategia de elaboración de una genoteca es bastante simple desde el punto de vista conceptual: como no se conoce el fragmento concreto que incluye el gen que se quiere clonar, se utilizan todos en los pasos siguientes. Con ello se obtiene una gran conjutno de poblaciones de organismos transformados con diferentes fragmentos de ADN, cada una de las cuales expresa un fragmento distinto del ADN manipulado. Es decir, tenemos una colección de genes procedentes del organismo inicial, lo que llamamos una genoteca. La población que exprese y produzca la proteína que interesa conseguir será la que utilizaremos más adelante. La ventaja de esta técnica es que, en un solo proceso, se pueden conseguir organismos transformados que expresen muchos genes, algunos de los cuales pueden ser interesantes. El inconveniente más importante es que es necesario manejar un gran número de poblaciones de organismos, lo que hace el proceso muy complejo y costoso.
La otra estrategia es el uso de sondas para identificar el fragmento de ADN que contiene el gen. Una sonda es un fragmento de un ácido nucleico (ADN o ARN) que se marca para poder reconocerla y que se une específicamente, gracias al principio de complementariedad de bases, a una región específica del ADN. Para identificar un gen a partir de una mezcla de fragmentos de ADN es posible, actualmente, sintetizar en el laboratorio una sonda de ADN, si se conoce la secuencia de bases o incluso la de aminoácidos de la proteína. Esta sonda, convenientemente marcada, se une al fragmento de ADN con una secuencia complementaria, lo que permite identificar esa secuencia en una electroforesis. A partir de ese momento se tiene el ADN concreto que se buscaba para seguir el proceso de clonación.
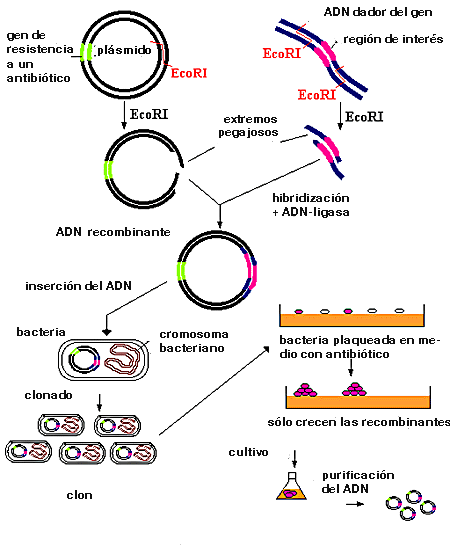
El objetivo final del proceso de clonación, no hay que olvidarlo, es introducir el gen del primer organismo en otro distinto. Para llevar a cabo este proceso hay que superar varios obstáculos, el primero de los cuales es que las células no aceptan sin más ADN extraño en su interior, sino que tratan de degradarlo. No obstante, hay algunos casos en los que sí se produce incorporación funcional de ADN en un organismo (los plásmidos en el caso de las bacterias, las infecciones víricas en todos los tipos celulares). Estos elementos capaces de incorporar ADN a las células que se van a transformar reciben el nombre de vectores, porque "transportan" el fragmento genético clonado hasta el interior de la célula.
Los vectores más utilizados en la clonación de genes son pues plásmidos o virus que no producen daños en la célula a la que atacan. Los biólogos moleculares que trabajan en este campo se han dedicado, durante mucho tiempo, a "manipular" estos vectores para introducir en ellos las características que resultan interesantes para su uso en la ingeniería genética. Tales características son:
- Poseer un "gen marcador", que codifique para una característica fácilmente reconocible. Esto permite identificar a las células que han absorbido y expresado el vector y por tanto el gen clonado.
- Tener secuencias de corte únicas para las enzimas de restricción que se utilizan en el proceso de clonación. La mayoría de los vectores habituales han sido manipulados hasta tal punto que todos ellos poseen puntos de corte únicos para multitud de restrictasas, localizados en zonas fuera de los genes necesarios para el funcionamiento del vector, de modo que se pueden utilizar en la mayor parte de los procedimientos de clonación sea cual sea la enzima apropiada para cortar el gen.

El vector se corta con la misma restrictasa que el material genético aislado que contiene el gen. Ambas muestras de ADN se mezclan, con lo que los extremos cohesivos de los dos fragmentos pueden asociarse entre sí gracias a la complementariedad de bases. Los fragmentos asociados así se unen covalentemente con ayuda de la ADN ligasa, de forma que, al final, se posee un "vector recombinante", una molécula que incluye el material genético del vector unido al fragmento de ADN que se pretende clonar.
El segundo obstáculo a superar es la propia entrada del material genético en la célula, lo que se conoce como transformación. Para que esto ocurra, hay que hacer que el ADN atraviese, al menos, la membrana celular. El proceso de transformación supone, en general, eliminar la pared celular si está presente y facilitar la formación de "poros" temporales en la membrana de las células, que deben cerrarse una vez que el vector haya tenido la posibilidad de entrar en la célula. Existe una estrategia alternativa, que consiste en utilizar una "pistola de genes", un aparato que dispara el ADN transformante, situado en la superficie de micropartículas de oro, contra las células a transformar.

No todas las células que son sometidas a la transformación captan el ADN exógeno. Los genes marcadores de los vectores sirven, en este paso del proceso, para seleccionar las células que sí lo han hecho, utilizando la expresión de dicho gen. Por ejemplo, si el marcador es un gen de resistencia a un antibiótico (como la ampicilina, que se utiliza en ingeniería genética porque no tiene valor clínico), se cultiva las células en presencia de esa sustancia; las que son capaces de crecer incluyen el plásmido.
De entre las células transformadas, finalmente, se selecciona aquellas que son capaces de producir la proteína de interés, de modo que se ha conseguido expresar el gen en un organismo diferente.
La clonación de genes suele utilizarse para producir proteínas de interés en microorganismos fáciles de cultivar en condiciones industriales. Por esa razón suelen utilizarse para estos fines bacterias, cuya manipulación es muy sencilla, o también levaduras (como
Saccharomyces cerevisiae) y hongos filamentosos (como los de los géneros
Aspergillus o
Penicillium) que, además de ser fáciles de cultivar, tienen la ventaja de ser eucariotas, con lo que introducen en las proteínas las mismas modificaciones postraduccionales que los organismos de los que proceden, y de secretar las proteínas con gran facilidad, lo que hace más sencilla la recuperacion del producto.
Pruebas de ADN
Las pruebas de ADN son procedimientos en los que se compara una muestra "problema" de ADN, cuya identidad se desconoce, con otra conocida, con el propósito de identificarlas. Las técnicas y herramientas concretas que se utilizarán dependen, en buena medida, del objetivo de la prueba, que puede ser relativamente diferente entre unos casos y otros. Entre los usos más habituales de este tipo de técnicas destacan:
- Identificación forense: el ADN de cada individuo es diferente al de los demás (excepto, claro, en el caso de gemelos idénticos), de modo que si se dispone de una muestra de ADN de identidad conocida, puede emplearse una pequeña cantidad de ADN para identificar, sin lugar a dudas, la identidad del donante problema. Este tipo de pruebas suelen realizarse con propósitos legales, especialmente en la identificación de autores de delitos, o de víctimas desconocidas.
- Pruebas de parentesco: de modo similar, el ADN de cada individuo comparte un cierto grado de similitud con el de sus parientes (por ejemplo, padres e hijos comparten un 50% de su material genético), de modo que pueden compararse muestras de dos individuos diferentes en busca de parecidos o diferencias. Entre este tipo de pruebas se incluyen, por ejemplo, las de determinación de paternidad: comparando la muestra del individuo con las de sus presuntos padres, puede llegar a determinarse la paternidad con un grado de fiabilidad muchísimo mayor que utilizando las pruebas genéticas tradicionales. Una extensión de este tipo de pruebas es la identificación de relaciones filogenéticas: puede compararse el ADN de especies diferentes para analizar el grado de similitud entre ellas, que está estrechamente relacionado con su parentesco evolutivo.
- Detección de enfermedades genéticas: la identificación de las versiones "normales" y mutadas de los genes que provocan enfermedades congénitas ayuda al diagnóstico precoz de este tipo de enfermedades, utilizando los genes mutados como sonda para la localización de dicha secuencia en los individuos que se quieren estudiar. El diagnóstico precoz es, en muchos casos, un factor importante en la mejora de la calidad de vida de los pacientes de este tipo de enfermedades.
En todo caso, las pruebas de ADN requieren disponer de una cantidad suficiente de muestra para poder ser comparada. La técnica conocida como PCR (reacción en cadena de la polimerasa) ha facilitado en gran medida este proceso, porque proporciona cantidades considerables de ADN a partir de una cantidad muy reducida de muestra.
La reacción en cadena de la polimerasa, desarrollada en los años 80' del siglo XX, se basa en la resistencia a la temperatura de la polimerasa de ADN procedente de un organismo termófilo, Tetrahymena thermophila, que se encuentra en las surgencias oceánicas. Los organismos que viven en esos ambientes son capaces de soportar temperaturas extremadamente elevadas, lo que supone que sus proteínas son especialmente resistentes a la desnaturalización por calor. Sabido esto, el fundamento de la técnica de PCR es bastante sencillo: la polimerasa es capaz de copiar moléculas de ADN monocatenario si dispone de un molde y nucleótidos trifosfato, y el ADN bicatenario se separa en sus dos hebras al aumentar la temperatura. Es decir, si se dispone de una cierta cantidad de ADN, se puede calentar para desnaturalizarlo (separarlo en sus hebras). Al añadirle polimerasa y nucleótidos trifosfato, la enzima copia las hebras de ADN. Si al finalizar el proceso se vuelve a calentar, se consigue la separación de las hebras, que pueden volver a servir de molde para un nuevo ciclo de replicación. Entre tanto, la enzima permanece activa gracias a su resistencia a la desnaturalización.
Así pues, partiendo de una pequeña muestra de ADN, fácil de obtener a partir de casi cualquier tejido biológico, se utiliza la PCR para amplificarla hasta conseguir la cantidad necesaria para realizar la prueba. Si se trata de comparar la muestra problema con un patrón conocido, ambas se cortan con una mezcla de restrictasas que proporcionan un conjunto de fragmentos de diferente tamaño, los cuales, a continuación, se someten a una electroforesis en gel. Se comparan entre sí los fragmentos del "patrón" y del "problema", lo que proporciona la información buscada.
- Si se trata de identificar a un individuo, los fragmentos de la muestra patrón y de la muestra problema deben ser idénticos.
- Si se trata de un análisis de parentesco, es necesario introducir en el estudio muestras de parientes conocidos del individuo (por ejemplo, en el caso de la determinación de paternidad es necesario incluir el ADN materno, para reconocer qué fragmentos proceden de la madre y cuáles pueden proceder del presunto padre).
En el caso de la detección de enfermedades genéticas, es necesario contar con una sonda de ADN con la secuencia específica que determina la enfermedad. La sonda se marca para poder reconocerla, y se añade a la muestra de ADN del paciente. La mezcla se calienta, para desnaturalizar el ADN, y se busca la presencia de material genético marcado. En caso de que aparezca, significa que está presente el alelo responsable de la enfermedad.
Clonación de individuos completos
La clonación de organismos pretende obtener individuos genéticamente idénticos a otros que ya existían. En general, en el caso de las plantas esto no supone ningún tipo de problema, ya que incluso es un proceso natural: la reproducción vegetativa (por esquejes). En el caso de los animales, también existen casos de "clonación natural": los gemelos idénticos, que se forman a partir de un único cigoto que, durante el proceso de división celular normal, se divide en varios. En algunas ocasiones, cada una de las partes puede llegar a desarrollarse por separado, con lo que se obtiene individuos que son totalmente idénticos entre sí.
El interés de la clonación, dejando aparte argumentos de ciencia ficción, está, por una parte, en obtener individuos animales que posean las mismas características genéticas que uno dado (por ejemplo ejemplares de ganado). Otra utilidad puede ser la de recuperar especies extinguidas a partir de su material genético conservado, como se ha tratado de hacer en la Universidad de Zaragoza con el sarrio.
La clonación, en si, es teóricamente sencilla, ya que consiste en sustituir el núcleo de un cigoto por el de una célula somática del organismo que se quiere clonar. Técnicamente, este procedimiento no plantea demasiados problemas; las células son lo suficientemente grandes como para poder ser visualizadas con el microscopio óptico, y se dispone de instrumental adecuado para manipular los núcleos. La célula se inmoviliza con una pipeta que absorbe el aire, haciendo ventosa, y el núcleo se extrae con una microjeringa. Si se trata del óvulo, el núcleo (que solo posee n cromosomas) se desecha. En cambio, si se trata de la célula donante, el núcleo se conserva dentro de la microjeringa y se utiliza para introducirlo en el ovocito.
El mayor problema de la clonación de animales, especialmente de animales complejos, está en conseguir que las células somáticas adultas se desdiferencien, lo que se consigue manteniéndolas en un medio de cultivo pobre en nutrientes.
 En realidad éste es un paso que ya se ha iniciado: la transformación genética ha permitido introducir genes de unos organismos en otros , en particular en microorganismos y plantas, que les permiten desarrollar nuevas funciones. Este es, sin duda, el primer paso hacia el diseño inteligente de organismos, y se dio hace ya bastante tiempo. Sin embargo, al menos por el momento, estos genes extraños no proporcionan a los organismos modificados ventajas adaptativas (menos mal). Lo único que hemos hecho hasta ahora ha sido manipular a los organismos para que nosotros podamos obtener un mayor rendimiento de su utilización.
En realidad éste es un paso que ya se ha iniciado: la transformación genética ha permitido introducir genes de unos organismos en otros , en particular en microorganismos y plantas, que les permiten desarrollar nuevas funciones. Este es, sin duda, el primer paso hacia el diseño inteligente de organismos, y se dio hace ya bastante tiempo. Sin embargo, al menos por el momento, estos genes extraños no proporcionan a los organismos modificados ventajas adaptativas (menos mal). Lo único que hemos hecho hasta ahora ha sido manipular a los organismos para que nosotros podamos obtener un mayor rendimiento de su utilización.