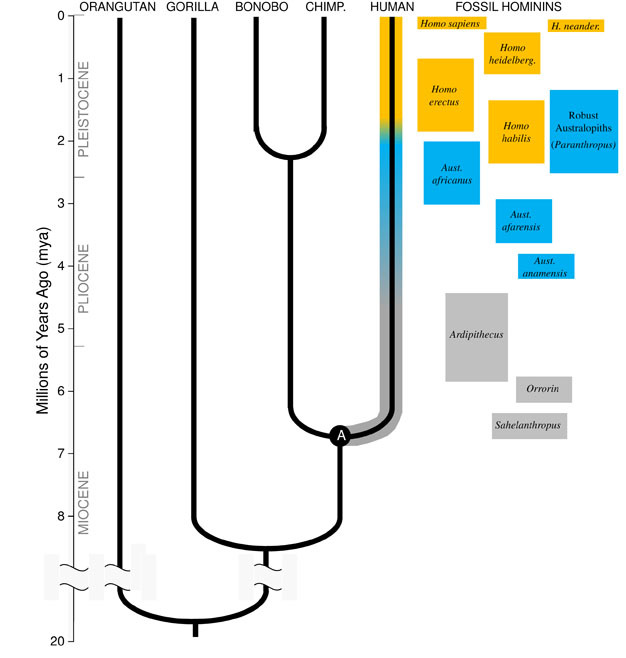
Darwin consiguió explicar el mecanismo a través del cual se produce la evolución biológica, la selección natural, pero no consiguió detallar ni la base sobre la que actúa la selección (la causa de la variabilidad genética de los organismos) ni el fundamento biológico de su teoría, es decir, qué determina que unas características de los individuos se transmitan de generación en generación, y por tanto tengan influencia evolutiva, mientras que otras no lo hacen.
Estos "pequeños detalles" sin explicar hicieron que, a medida que se fue desarrollando la Genética, con el redescubrimiento de las leyes de Mendel y el desarrollo de la teoría cromosómica de la herencia, la teoría evolutiva fuera resultando cada vez más "incómoda", en el sentido de que, aunque permitiera explicar una gran cantidad de fenómenos biológicos, no encajaba bien con las nuevas explicaciones derivadas de otras teorías emergentes e igualmente explicativas.
Además, los dos detalles se explicaban bastante bien con ayuda de la teoría cromosómica de la herencia. La variabilidad genética se produce por medio de las mutaciones, que alteran la información genética almacenada en el ADN, y se extienden por la población mediante la reproducción sexual y las migraciones. Asimismo, la reordenación cromosómica y la recombinación que tienen lugar durante la meiosis contribuyen a generar nuevos fenotipos, gracias a que permiten mezclar entre sí en un número potencialmente infinito de formas las diferentes variantes producidas por mutación.
Sin embargo, estas variaciones son de naturaleza "cuántica": si una mutación da lugar a una proteína diferente a la original, los individuos de la población llevarán la primera forma de la proteína o la segunda, pero no una forma intermedia entre ellas, mientras que la teoría evolutiva de Darwin defendía el carácter continuo del cambio...
Así pues, las leyes de Mendel y la teoría cromosómica de Morgan, que demostraban el caracter "cuantizado" de las características hereditarias parecían oponerse frontalmente a la idea del cambio gradual sostenida por Darwin. El primer avance hacia la unión de las dos visiones del mundo biológico se produjo cuando Fisher consiguió establecer un modelo matemático que demostraba que la variación continua en las características de los individuos podía ser el resultado de cambios "discretos" (es decir, no continuos) en muchos genes diferentes.
El avance definitivo hacia la nueva teoría se produjo cuando se pudo relacionar la teoría evolutiva con la genética de poblaciones. La teoría de la selección natural habla de la persistencia de unos individuos frente a otros, como consecuencia de sus diferentes características, en el seno de una población. Para poder pasar de un principio a otro es necesario saber algo de la población en la que se encuentran los individuos que van a sufrir la selección natural y cuyas características genéticas van a ser transmitidas o eliminadas.
El problema de la especiación
 La teoría darwiniana no plantea demasiados problemas para explicar la evolución filética (o microevolución): los cambios sufridos por una población a lo largo del tiempo, como resultado de una presión selectiva permanente. Simplemente, la acumulación de pequeñas variaciones provocadas por mutaciones da lugar a un conjunto de cambios graduales, de acuerdo con el modelo propuesto por Fisher. Los individuos de cada nueva generación podían ser ligeramente distintos a sus progenitores, con modificaciones casi imperceptibles, pero al cabo de un periodo de tiempo suficientemente largo se habrán acumulado tantas diferencias que los individuos finales y sus remotos antepasados pueden llegar a ser casi totalmente diferentes.
La teoría darwiniana no plantea demasiados problemas para explicar la evolución filética (o microevolución): los cambios sufridos por una población a lo largo del tiempo, como resultado de una presión selectiva permanente. Simplemente, la acumulación de pequeñas variaciones provocadas por mutaciones da lugar a un conjunto de cambios graduales, de acuerdo con el modelo propuesto por Fisher. Los individuos de cada nueva generación podían ser ligeramente distintos a sus progenitores, con modificaciones casi imperceptibles, pero al cabo de un periodo de tiempo suficientemente largo se habrán acumulado tantas diferencias que los individuos finales y sus remotos antepasados pueden llegar a ser casi totalmente diferentes.
Sin embargo, la especiación o macroevolución, es decir, la aparición de especies distintas a partir de una sola población, resulta mucho más difícil de explicar mediante cambios graduales. Por una parte, estos cambios deberían poder observarse en el registro fósil como una serie de formas intermedias entre una especie y la otra. En realidad esto no ocurre nunca o casi nunca, en parte debido al hecho de que la fosilización es un fenómeno muy poco frecuente, y la conservación y hallazgo de los fósiles aún más. Esta falta de los "eslabones perdidos" ha sido utilizada frecuentemente como argumento en contra de la evolución bioológica.
Pero hay otro motivo que hace difícil de explicar la especiación mediante cambios graduales: si dos individuos se diferencian ligeramente, deberían poder cruzarse entre sí y tener descendencia fértil, de modo que la población estaría formada por híbridos entre ellos. De ese modo resulta complicado poder explicar cómo se acumulan suficientes diferencias como para separar las nuevas especies.
 Una explicación sencilla al proceso de especiación es suponer que una población queda separada en dos partes debido a la existencia de una barrera geográfica que impide el contacto entre los dos grupos, por ejemplo un río, un brazo de mar o una cordillera montañosa. Con el paso del tiempo las dos poblaciones van sufriendo mutaciones y procesos de selección natural que pueden ser diferentes, puesto que ocupan ambientes distintos. Si el proceso continúa durante el tiempo suficiente las diferencias serán tan grandes que los dos grupos se habrán convertido en especies distintas, sin posibilidad de cruzarse entre ellas. Este modelo de especiación se denomina modelo alopátrido (de alo = otro, distinto) porque supone que la diversificación tiene lugar en espacios geográficos diferentes, y parece ser el más proceso más frecuente a lo largo de la historia evolutiva.
Una explicación sencilla al proceso de especiación es suponer que una población queda separada en dos partes debido a la existencia de una barrera geográfica que impide el contacto entre los dos grupos, por ejemplo un río, un brazo de mar o una cordillera montañosa. Con el paso del tiempo las dos poblaciones van sufriendo mutaciones y procesos de selección natural que pueden ser diferentes, puesto que ocupan ambientes distintos. Si el proceso continúa durante el tiempo suficiente las diferencias serán tan grandes que los dos grupos se habrán convertido en especies distintas, sin posibilidad de cruzarse entre ellas. Este modelo de especiación se denomina modelo alopátrido (de alo = otro, distinto) porque supone que la diversificación tiene lugar en espacios geográficos diferentes, y parece ser el más proceso más frecuente a lo largo de la historia evolutiva.
Se han propuesto también modelos de especiación simpátrida, es decir, que tienen lugar en un mismo ecosistema, aunque las condiciones necesarias para que se de son bastante específicas y difíciles de cumplir.
Algunas heterodoxias evolutivas
Cuando una teoría se impone dentro de una comunidad científica, es bastante habitual que sus defensores "cierren filas" en torno a ella, rechazando con fuerza las matizaciones que van descubriéndose con el paso del tiempo. Finalmente, algunas de esas matizaciones acaban por incorporarse a la teoría, enriqueciéndola, no sin que antes se haya producido un intenso debate que muchas veces ayuda al progreso y al perfeccionamiento de la teoría original.
El caso de la evolución es bastante parecido al de otros sistemas teóricos, tanto en el área de la Física como en el de la Geología, por ejemplo. Los principales debates, que aún siguen manteniéndose, tienen que ver con los mecanismos con los que opera la evolución y con su ritmo.
Los mecanismos de la evolución: ¿solo selección natural?
Cuando escribió el "Origen de las Especies" el propio Darwin se encargó de señalar su convencimiento de que la selección natural era el principal mecanismo de la evolución, pero no el único. Sin embargo, cuando se desarrolló la teoría evolutiva moderna, el Neodarwinismo, sus defensores afirmaron tajantemente que toda la evolución se produce siempre mediante selección natural, por lo que desde entonces los biólogos evolucionistas se han dedicado a buscar las presiones evolutivas que expliquen cada una de las características de los diferentes organismos.
Sin embargo, hay muchos evolucionistas que defienden la posibilidad de que, en algunos casos, la evolución no se deba exclusivamente a la selección natural, sino a la "deriva genética", es decir, a procesos aleatorios.
 Se han descrito varios mecanismos que pueden ejemplificar la evolución de una población mediante deriva genética. Uno de ellos es el efecto de "cuello de botella": si una población sufre una reducción drástica en su número de individuos como consecuencia de una catástrofe accidental (una inundación, un terremoto, una erupción volcánica...) en la que no ocurre selección natural, es decir, todos los individuos tienen probabilidades parecidas de morir. La población más pequeña es una muestra aleatoria de la original, y los tipos de individuos que la forman no tienen por qué representar las proporciones que existían inicialmente, por lo que la nueva población tendrá características diferentes, que no se pueden atribuir a la selección natural.
Se han descrito varios mecanismos que pueden ejemplificar la evolución de una población mediante deriva genética. Uno de ellos es el efecto de "cuello de botella": si una población sufre una reducción drástica en su número de individuos como consecuencia de una catástrofe accidental (una inundación, un terremoto, una erupción volcánica...) en la que no ocurre selección natural, es decir, todos los individuos tienen probabilidades parecidas de morir. La población más pequeña es una muestra aleatoria de la original, y los tipos de individuos que la forman no tienen por qué representar las proporciones que existían inicialmente, por lo que la nueva población tendrá características diferentes, que no se pueden atribuir a la selección natural.
 Otro caso que puede explicar la evolución sin selección es el "efecto fundador", que se produce cuando un grupo de individuos de una población coloniza un hábitat nuevo. En este caso los individuos que migran no tienen por qué representar la proporción de los individuos presentes en la población original (en realidad, es un caso muy parecido al efecto cuello de botella). De nuevo, el "muestreo" de una población grande puede alterar la proporción de individuos de la población resultante.
Otro caso que puede explicar la evolución sin selección es el "efecto fundador", que se produce cuando un grupo de individuos de una población coloniza un hábitat nuevo. En este caso los individuos que migran no tienen por qué representar la proporción de los individuos presentes en la población original (en realidad, es un caso muy parecido al efecto cuello de botella). De nuevo, el "muestreo" de una población grande puede alterar la proporción de individuos de la población resultante.
El ritmo del cambio
Una cuestión más que provoca la discusión entre los evolucionistas es el ritmo con el que se produce el cambio evolutivo, y más concretamente la discusión acerca de si se produce de un modo más o menos constante (gradualismo) o de si, en determinadas circunstancias, se produce una aceleración de la tasa de cambio (equilibrio puntuado).
 Los defensores de esta hipótesis sostienen que los procesos evolutivos pueden ser lentos durante largos periodos de tiempo, especialmente si durante ellos las condiciones ambientales son muy estables. En estas fases la evolución sería básicamente gradual (microevolución), pero en ciertos momentos, posiblemente relacionados con cambios ambientales bruscos, podría producirse un aumento considerable de la tasa de cambio, que daría lugar a la rápida aparición de nuevas especies.
Los defensores de esta hipótesis sostienen que los procesos evolutivos pueden ser lentos durante largos periodos de tiempo, especialmente si durante ellos las condiciones ambientales son muy estables. En estas fases la evolución sería básicamente gradual (microevolución), pero en ciertos momentos, posiblemente relacionados con cambios ambientales bruscos, podría producirse un aumento considerable de la tasa de cambio, que daría lugar a la rápida aparición de nuevas especies.
Existen mecanismos genéticos que pueden explicar los cambios evolutivos bruscos, como las mutaciones cromosómicas (pérdida o duplicación de fragmentos de cromosoma, fusión de cromosomas) o genómicas (poliploidía, es decir, multiplicación del número de cromosomas porque no se separan durante la división celular). Todos esos procesos tienen en común que impiden de forma inmediata el apareamiento entre individuos "silvestres" y mutantes, con lo que crean una barrera reproductiva entre ellos. La poliploidía, en especial, facilita además la mutación de los genes y por lo tanto la acumulación de variedades, ya que como los individuos tienen duplicados todos los genes, una de las copias puede mutar sin provocar efectos deletéreos en el organismo, que conserva el otro gen intacto.
Genética del individuo y de la población
 Desde el punto de vista de la genética describimos a los individuos por medio de su "composición alélica". Un individuo puede ser homocigoto (dominante o recesivo) o heterocigoto. Para describir en términos genéticos una población necesitamos saber el número (mejor aún, la frecuencia) de individuos de cada tipo que la forman.
Desde el punto de vista de la genética describimos a los individuos por medio de su "composición alélica". Un individuo puede ser homocigoto (dominante o recesivo) o heterocigoto. Para describir en términos genéticos una población necesitamos saber el número (mejor aún, la frecuencia) de individuos de cada tipo que la forman.
Igual que ocurre en la genética individual, en la genética de poblaciones es importante conocer no solo los fenotipos, sino también los genotipos de los individuos, porque de ellos dependen las características que se transmitirán a la siguiente generación. Igual que ocurre en la segunda ley de Mendel, los alelos recesivos pueden estar "ocultos" en los heterocigotos, pero van a ser transmitidos y "aflorarán" en la siguiente generación. Esto tiene importancia desde el punto de vista evolutivo: un individuo recesivo puede tener una probabilidad de supervivencia diferente a la de sus hermanos dominantes.
Así que al analizar genéticamente una población no solo nos interesa conocer la frecuencia de los fenotipos, sino también la de los alelos. Pero, otra vez igual que lo que sucede con los individuos, tenemos el problema de que no podemos distinguir los individuos homocigóticos dominantes de los heterocigóticos. Sin embargo, sí que podemos calcular su frecuencia utilizando un modelo teórico, la ley de Hardy Weimberg.
Podemos estudiar la genética de poblaciones de un modo simple analizando un gen con dos alelos (igual que en las leyes de Mendel) en una población en equilibrio (en la que no cambian las frecuencias génicas), aplicando el cuadrado de Punnett. En este caso, introducimos en él la probabilidad de que cada individuo posea el alelo dominante (A) o el recesivo (a), y obtenemos lo siguiente:
Además, los dos detalles se explicaban bastante bien con ayuda de la teoría cromosómica de la herencia. La variabilidad genética se produce por medio de las mutaciones, que alteran la información genética almacenada en el ADN, y se extienden por la población mediante la reproducción sexual y las migraciones. Asimismo, la reordenación cromosómica y la recombinación que tienen lugar durante la meiosis contribuyen a generar nuevos fenotipos, gracias a que permiten mezclar entre sí en un número potencialmente infinito de formas las diferentes variantes producidas por mutación.
Sin embargo, estas variaciones son de naturaleza "cuántica": si una mutación da lugar a una proteína diferente a la original, los individuos de la población llevarán la primera forma de la proteína o la segunda, pero no una forma intermedia entre ellas, mientras que la teoría evolutiva de Darwin defendía el carácter continuo del cambio...
Así pues, las leyes de Mendel y la teoría cromosómica de Morgan, que demostraban el caracter "cuantizado" de las características hereditarias parecían oponerse frontalmente a la idea del cambio gradual sostenida por Darwin. El primer avance hacia la unión de las dos visiones del mundo biológico se produjo cuando Fisher consiguió establecer un modelo matemático que demostraba que la variación continua en las características de los individuos podía ser el resultado de cambios "discretos" (es decir, no continuos) en muchos genes diferentes.
El avance definitivo hacia la nueva teoría se produjo cuando se pudo relacionar la teoría evolutiva con la genética de poblaciones. La teoría de la selección natural habla de la persistencia de unos individuos frente a otros, como consecuencia de sus diferentes características, en el seno de una población. Para poder pasar de un principio a otro es necesario saber algo de la población en la que se encuentran los individuos que van a sufrir la selección natural y cuyas características genéticas van a ser transmitidas o eliminadas.
El problema de la especiación
Sin embargo, la especiación o macroevolución, es decir, la aparición de especies distintas a partir de una sola población, resulta mucho más difícil de explicar mediante cambios graduales. Por una parte, estos cambios deberían poder observarse en el registro fósil como una serie de formas intermedias entre una especie y la otra. En realidad esto no ocurre nunca o casi nunca, en parte debido al hecho de que la fosilización es un fenómeno muy poco frecuente, y la conservación y hallazgo de los fósiles aún más. Esta falta de los "eslabones perdidos" ha sido utilizada frecuentemente como argumento en contra de la evolución bioológica.
Pero hay otro motivo que hace difícil de explicar la especiación mediante cambios graduales: si dos individuos se diferencian ligeramente, deberían poder cruzarse entre sí y tener descendencia fértil, de modo que la población estaría formada por híbridos entre ellos. De ese modo resulta complicado poder explicar cómo se acumulan suficientes diferencias como para separar las nuevas especies.
Se han propuesto también modelos de especiación simpátrida, es decir, que tienen lugar en un mismo ecosistema, aunque las condiciones necesarias para que se de son bastante específicas y difíciles de cumplir.
Algunas heterodoxias evolutivas
Cuando una teoría se impone dentro de una comunidad científica, es bastante habitual que sus defensores "cierren filas" en torno a ella, rechazando con fuerza las matizaciones que van descubriéndose con el paso del tiempo. Finalmente, algunas de esas matizaciones acaban por incorporarse a la teoría, enriqueciéndola, no sin que antes se haya producido un intenso debate que muchas veces ayuda al progreso y al perfeccionamiento de la teoría original.
El caso de la evolución es bastante parecido al de otros sistemas teóricos, tanto en el área de la Física como en el de la Geología, por ejemplo. Los principales debates, que aún siguen manteniéndose, tienen que ver con los mecanismos con los que opera la evolución y con su ritmo.
Los mecanismos de la evolución: ¿solo selección natural?
Cuando escribió el "Origen de las Especies" el propio Darwin se encargó de señalar su convencimiento de que la selección natural era el principal mecanismo de la evolución, pero no el único. Sin embargo, cuando se desarrolló la teoría evolutiva moderna, el Neodarwinismo, sus defensores afirmaron tajantemente que toda la evolución se produce siempre mediante selección natural, por lo que desde entonces los biólogos evolucionistas se han dedicado a buscar las presiones evolutivas que expliquen cada una de las características de los diferentes organismos.
Sin embargo, hay muchos evolucionistas que defienden la posibilidad de que, en algunos casos, la evolución no se deba exclusivamente a la selección natural, sino a la "deriva genética", es decir, a procesos aleatorios.
El ritmo del cambio
Una cuestión más que provoca la discusión entre los evolucionistas es el ritmo con el que se produce el cambio evolutivo, y más concretamente la discusión acerca de si se produce de un modo más o menos constante (gradualismo) o de si, en determinadas circunstancias, se produce una aceleración de la tasa de cambio (equilibrio puntuado).
Existen mecanismos genéticos que pueden explicar los cambios evolutivos bruscos, como las mutaciones cromosómicas (pérdida o duplicación de fragmentos de cromosoma, fusión de cromosomas) o genómicas (poliploidía, es decir, multiplicación del número de cromosomas porque no se separan durante la división celular). Todos esos procesos tienen en común que impiden de forma inmediata el apareamiento entre individuos "silvestres" y mutantes, con lo que crean una barrera reproductiva entre ellos. La poliploidía, en especial, facilita además la mutación de los genes y por lo tanto la acumulación de variedades, ya que como los individuos tienen duplicados todos los genes, una de las copias puede mutar sin provocar efectos deletéreos en el organismo, que conserva el otro gen intacto.
Genética del individuo y de la población
Igual que ocurre en la genética individual, en la genética de poblaciones es importante conocer no solo los fenotipos, sino también los genotipos de los individuos, porque de ellos dependen las características que se transmitirán a la siguiente generación. Igual que ocurre en la segunda ley de Mendel, los alelos recesivos pueden estar "ocultos" en los heterocigotos, pero van a ser transmitidos y "aflorarán" en la siguiente generación. Esto tiene importancia desde el punto de vista evolutivo: un individuo recesivo puede tener una probabilidad de supervivencia diferente a la de sus hermanos dominantes.
Así que al analizar genéticamente una población no solo nos interesa conocer la frecuencia de los fenotipos, sino también la de los alelos. Pero, otra vez igual que lo que sucede con los individuos, tenemos el problema de que no podemos distinguir los individuos homocigóticos dominantes de los heterocigóticos. Sin embargo, sí que podemos calcular su frecuencia utilizando un modelo teórico, la ley de Hardy Weimberg.
Podemos estudiar la genética de poblaciones de un modo simple analizando un gen con dos alelos (igual que en las leyes de Mendel) en una población en equilibrio (en la que no cambian las frecuencias génicas), aplicando el cuadrado de Punnett. En este caso, introducimos en él la probabilidad de que cada individuo posea el alelo dominante (A) o el recesivo (a), y obtenemos lo siguiente:
| A (p) | a (q) | |
| A (p) | AA (p2) | Aa (pq) |
| a (q) | Aa (pq) | aa (q2) |
Es decir, si suponemos que las frecuencias de los alelos son p para el alelo dominante (A) y q para el recesivo (a), la frecuencia en la población de cada genotipo será la siguiente:
- Frecuencia del genotipo homocigoto dominante p2
- Frecuencia del genotipo homocigoto recesivo q2
- Frecuencia del genotipo heterocigoto 2pq
Se puede calcular fácilmente la frecuencia del alelo dominante porque, como el gen solo tiene dos alelos, la suma de sus frecuencias es la unidad:
p = 1 - q.
El siguiente paso: la selección natural
Ahora ya tenemos descrita la población desde el punto de vista genético. Sabemos cuántos individuos son homocigotos y cuántos son heterocigotos, de modo que podemos calcular el número de individuos de cada tipo que habrá en la siguiente generación.
La selección natural actúa contra los fenotipos, contra las características que se manifiestan en la población, provocando que los individuos que poseen una característica poco adaptativa tengan más posibilidades de morir que otros. Hablando en términos genéticos, lo que hace es que la frecuencia de un determinado fenotipo se reduzca frente a la de otro u otros. Esto tiene consecuencia en las frecuencias alélicas, que también cambian, de modo que la siguiente generación va a tener un número de individuos distinto para cada fenotipo. Si esa mortalidad diferente (selección) se mantiene a lo largo del tiempo (presión selectiva), al cabo de unas cuantas generaciones las frecuencias fenotípicas, y las alélicas, habrán variado considerablemente, siguiendo una cierta tendencia. La población final será, por tanto, distinta a la original. Hemos llegado a la evolución.
La ley de Hardy Weimberg nos da una imagen de una población "congelada", en la que el número de individuos de cada tipo es el mismo a lo largo del tiempo, pero podemos utilizarla para elaborar un modelo matemático sencillo de la selección a favor o en contra de uno de los alelos. Se trata, simplemente, de introducir la mortalidad diferencial de cada genotipo y recalcular las frecuencias alélicas a partir de los individuos que sobreviven en cada generación. Repitiendo (iterando) estos datos durante un cierto periodo de tiempo se puede simular el comportamiento de la población.
Puedes encontrar un ejemplo de este tipo de modelos en este enlace. Puedes utilizarlo cambiando la frecuencia de uno de los alelos (la del segundo alelo depende de ella) o la mortalidad de los fenotipos, y te permitirá observar cómo van cambiando a lo largo del tiempo las frecuencias alélicas y las fenotípicas en los diferentes casos que se analizan: herencia con dominancia simple, codominancia y herencia ligada al sexo.
Puedes encontrar un ejemplo de este tipo de modelos en este enlace. Puedes utilizarlo cambiando la frecuencia de uno de los alelos (la del segundo alelo depende de ella) o la mortalidad de los fenotipos, y te permitirá observar cómo van cambiando a lo largo del tiempo las frecuencias alélicas y las fenotípicas en los diferentes casos que se analizan: herencia con dominancia simple, codominancia y herencia ligada al sexo.