
Un tejido es un grupo de células y de material circundante que trabajan en conjunto para cumplir una función determinada. En los metazoos (animales pluricelulares) existen cuatro tipos fundamentales de tejidos:
- El tejido epitelial reviste la superficie del cuerpo, tapiza los órganos huecos, cavidades y conductos del organismo y da origen a las glándulas.
- El tejido conectivo protege y da soporte al cuerpo, almacena energía y proporciona defensa frente a infecciones.
- El tejido muscular produce la fuerza necesaria para permitir el movimiento en el organismo.
- El tejido nervioso recibe información tanto del interior como del exterior del cuerpo y responde a ella generando impulsos eléctricos que la transmiten, contribuyendo a mantener la homeostasis.
Los tejidos epiteliales se caracterizan porque sus células están dispuestas en capas continuas, sin apenas separación entre ellas. El tejido puede tener una única capa de células (monoestratificado) o más de una (pluriestratificado). Las células que lo componen están estrechamente unidas entre sí mediante distintos tipos de estructuras especializadas, de forma que todos los epitelios tapizan superficies, es decir, recubren totalmente los tejidos subyacentes sin que ninguna molécula pueda atravesar el tejido pasando entre sus células. Debido a la gran proximidad entre las células que componen los epitelios la sustancia intercelular de este tipo de tejidos es escasa y juega muy poco papel en su funcionamiento.
Las células de los epitelios están polarizadas, lo que significa que sus superficies celulares tienen distintas funciones. En su parte "inferior", llamada más propiamente basal, todos los epitelios están separados del tejido conectivo que hay bajo ellos por una capa acelular llamada membrana basal, en la que se pueden distinguir dos partes: la lámina basal es segregada por el propio epitelio, mientras que la lámina reticular está producida por el tejido conectivo. Ambas están constituidas fundamentalmente por proteínas fibrosas, fundamentalmente colágeno.
En la diabetes mellitus que no se trata adecuadamente se produce un engrosamiento de la membrana basal de los capilares sanguíneos, especialmente en los ojos y los riñones, lo que puede dar lugar a ceguera y a insuficiencia renal.
Los vasos sanguíneos no atraviesan la membrana basal, de modo que los epitelios son "avasculares". Esto supone que la nutrición de las células se produce mediante difusión de los nutrientes a través de la membrana basal, lo que limita el espesor máximo de los epitelios. Sin embargo, los nervios sí que penetran en el tejido, de modo que pueden llegar a poseer una gran sensibilidad como ocurre, por ejemplo, en ciertas zonas de la epidermis.

Existen dos grandes tipos de tejidos epiteliales: los de revestimiento y los glandulares. Los primeros cubren la piel y algunos órganos internos, forman la capa más interna de los vasos sanguíneos y de los conductos y cavidades corporales y tapizan el interior de los aparatos respiratorio, digestivo, urinario y reproductor. Por su parte los epitelios glandulares constituyen la parte secretora de todas las glándulas del organismo.
Las glándulas son células (las hay unicelulares) o grupos de células que producen y liberan al exterior alguna sustancia útil. Esa secreción puede ser vertida hacia el exterior del cuerpo, o hacia cavidades o tubos que se abren al exterior, como el aparato digestivo. En ese caso se dice que las glándulas son exocrinas. Existen también glándulas cuyo producto de secreción va a parar al sistema cardiovascular, en cuyo caso se denominan endocrinas.
Los epitelios de revestimiento se clasifican según su disposición en capas y la forma de sus células. En cuanto a la disposición en capas se distinguen:
- Epitelios simples: están formados por una única capa de células, de modo que todas se apoyan directamente en la membrana basal y tienen sus núcleos aproximadamente a la misma altura cuando se observann al microscopio. En general se ocupan de la secreción y la absorción de sustancias.
- Epitelios pluriestratificados: las células se disponen en dos o más capas, de modo que se observan varias filas de núcleos a diferentes alturas. En general se encargan de proteger tejidos expuestos a daño o entre los que se produce rozamiento.
 Epitelios pseudoestratificados: los núcleos de las células se encuentran a diferentes alturas, y no todas las células alcanzan la misma altura, pero todas se apoyan en la membrana basal. Un ejemplo típico de esta clase de epitelios es el de la vejiga urinaria, cuya característica fundamental es que sus células cambian su forma, de modo que el epitelio tiene una forma "colapsada" en la que es más alto pero tiene poca superficie, y otra "distendida", en la que ocupa una superficie mayor, lo que permite que aumente el volumen de la vejiga a medida que se va llenando con la orina.
Epitelios pseudoestratificados: los núcleos de las células se encuentran a diferentes alturas, y no todas las células alcanzan la misma altura, pero todas se apoyan en la membrana basal. Un ejemplo típico de esta clase de epitelios es el de la vejiga urinaria, cuya característica fundamental es que sus células cambian su forma, de modo que el epitelio tiene una forma "colapsada" en la que es más alto pero tiene poca superficie, y otra "distendida", en la que ocupa una superficie mayor, lo que permite que aumente el volumen de la vejiga a medida que se va llenando con la orina.

En cuanto a la forma de las células, los epitelios de revestimiento se clasifican en:
- Epitelios pavimentosos, también llamados planos o escamosos: sus células son aplanadas, lo que facilita el intercambio de sustancias a su través.
- Epitelios cúbicos: sus células son aproximadamente isodiamétricas, es decir, su anchura es bastante similar a su altura. Suelen tener forma cúbica o hexagonal, y pueden presentar microvellosidades en su superficie apical (la opuesta a la que limita con la membrana basal), en cuyo caso pueden participar en procesos de absorción o de secreción.
- Epitelios cilíndricos o columnares: sus células son más altas que anchas. Suelen tener como función la protección de tejidos subyacentes, aunque también pueden tener microvellosidades, realizando entonces funciones de absorción.
- Los epitelios de transición tienen algunas células cúbicas, generalmente situadas en la parte basal del tejido y otras pavimentosas.

Las glándulas pueden clasificarse según el número de células que las forman y según su estructura, pero también teniendo en cuenta cuál es el destino de esas secreciones. Desde este punto de vista se distinguen las glándulas endocrinas, que vierten hacia el sistema cardiovascular, las paracrinas, que liberan sustancias que van a parar a otras zonas del cuerpo próximas a la propia glándula, sin necesidad de que sean transportadas por la sangre, y las exocrinas, que segregan al exterior del cuerpo. A su vez estas últimas se diferencian entre sí según el modo en el que producen la secreción:
- Las glándulas merocrinas sintetizan sus secreciones en los ribosomas y las liberan mediante exocitosis a través de vesículas del aparato de Golgi.
- Las glándulas apocrinas acumulan sus productos de secreción en la parte apical de las células. Esta parte se desprende y se repara, con lo que la célula sigue siendo funcional posteriormente.
- En las glándulas holocrinas el producto de secreción se acumula por todo el citoplasma y para ser liberado la célula debe destruirse.

Tejidos conectivos o conjuntivos

Los conectivos son una familia de tejidos que se caracterizan por tener una matriz extracelular muy abundante, formada por proteínas con forma de hilo, que reciben el nombre de fibras que se depositan en un sustrato llamado matriz fundamental.
La matriz extracelular es secretada por las células del tejido, y sus características, que pueden ser muy diferentes de unos conectivos a otros, son las que determinan las propiedades de cada tejido en particular.
Los tejidos conectivos no se encuentran nunca en la superficie del cuerpo y la mayoría de ellos (excepto cartílago y tendones) poseen vasos sanguíneos. Todos excepto los cartílagos están inervados.
Todos los tejidos conectivos proceden de una misma capa de células embrionarias, el mesodermo. A partir de esta capa se forman varios tipos de células no totalmente diferenciadas, que son las que dan origen a los tejidos conectivos.
Las células con capacidad de división que forman parte de los tejidos conectivos se identifican porque se denominan con el sufijo -blasto, que significa germen. Cuando maduran pierden su capacidad de reproducirse y de generar matriz. Para diferenciarlas se denominan con el sufijo -cito.
Los diferentes tipos de tejidos conectivos poseen distintos tipos celulares. Así, las células del tejido óseo que tienen capacidad de dividirse se denominan osteoblastos, mientras que cuando maduran se transforman en osteoclastos. En el cartílago, las células jóvenes son los condroblastos y las maduras se llaman condrocitos. En cambio, en los tejidos conectivos laxos y densos las células conservan permanentemente su capacidad de división y reciben el nombre de fibroblastos.

Los fibroblastos son células grandes y aplanadas, con prolongaciones citoplasmáticas ramificadas y un citoesqueleto muy bien desarrollado en su interior y que son capaces de moverse a lo largo del tejido. Se encuentran en diferentes tejidos conectivos, siendo el tipo celular más abundante en muchos de ellos, donde son los encargados de mantener y producir la matriz celular. De hecho, los fibroblastos actúan como verdaderas "fábricas químicas", sintetizando no solo los componentes de la matriz (sustancias solubles, polisacáridos como el ácido hialurónico o proteínas como el colágeno o la elastina), sino también, entre otras sustancias, factores de crecimiento o enzimas que son fundamentales para el funcionamiento correcto de los tejidos.
Además de los fibroblastos los tejidos conectivos presentan células especializadas, que se encuentran en sus respectivos tejidos. Así, las células que forman parte del cartílago son los condroblastos que se transforman en condrocitos, mientras que en el tejido óseo los osteoblastos se diferencian para dar lugar a osteocitos. Los adipocitos son células especializadas en el almacenamiento de grasas, concretamente de triacilglicéridos. Se encuentran especialmente por debajo de la piel y junto a algunos órganos internos como el corazón y cumplen una doble misión: servir como reserva energética y como aislante térmico.
En los tejidos conectivos se encuentran también varios tipos de leucocitos que pueden entrar y salir a estos tejidos procedentes del sistema cardiovascular para ocuparse de la defensa inmunitaria. Entre los distintos tipos de glóbulos blancos que llegan a los conectivos se incluyen los macrófagos, de forma irregular y con prolongaciones citoplasmáticas que les permiten fagocitar bacterias y restos celulares. Algunos están fijos en ciertos órganos (pulmón, bazo), mientras que otros recorren distintos tejidos dirigiéndose hacia las zonas en las que se producen procesos inflamatorios (circulantes). Además se pueden encontrar también mastocitos, que producen histamina, sustancia que interviene en los procesos de inflamación y que tienen capacidad fagocítica, así como linfocitos, que penetran desde el circulatorio en casos de infección, y células plasmáticas, que segregan anticuerpos. En su mayoría las células plasmáticas se encuentran en los tejidos conectivos, glándulas salivares, gánglios linfáticos y médula ósea.
Matriz extracelular
En los tejidos conectivos el espacio entre células es, en general, bastante amplio, y está ocupado por una matriz extracelular que determina las propiedades del sistema y juega importantes papeles en su funcionamiento.
La matriz extracelular de los tejidos conjuntivos está formada por dos componentes básicos: la sustancia fundamental, también llamada matriz amorfa, y las fibras, que son en realidad diferentes tipos de proteínas alargadas que le proporcionan distintas propiedades.

La sustancia amorfa une las células entre sí y les proporciona soporte mecánico, almacena agua y proporciona el medio de intercambio entre la sangre y las células, además de intervenir activamente en los procesos fisiológicos de las células. Contiene fundamentalmente agua y sustancias orgánicas de gran tamaño que reciben el nombre de proteoglicanos y que están constituidas por la unión de proteínas de adhesión y varios tipos de polisacáridos denominados conjuntamente glucosaminoglicanos. Los tipos más importantes de glucosaminoglicanos (o glucosaminoglucanos, ambos nombres son totalmente equivalentes) son el ácido hialurónico, que lubrica las células y las mantiene unidas entre sí, y el condroitín sulfato, que proporciona capacidad de soporte y adhesividad a cartílago, hueso, piel y vasos sanguíneos. Las proteínas de adhesión se encargan de unir la matriz amorfa con los componentes celulares del tejido.
Las fibras que componen la matriz extracelular de los tejidos conectivos son, en realidad, conjuntos de proteínas con estructura similar a hilos o cables que le proporcionan resistencia y elasticidad. Existen tres tipos de fibras en los tejidos conectivos: las fibras de colágeno, las elásticas y las reticulares.
 Todos los tipos de tejido conectivo adulto derivan de un único tipo de tejido conectivo que se forma durante el desarrollo embrionario. En el adulto este tejido se diversifica considerablemente, dando lugar a una gran variedad de tejidos diferentes. Una posible clasificación de los conectivos adultos es la que los diferencia según sus características mecánicas:
Todos los tipos de tejido conectivo adulto derivan de un único tipo de tejido conectivo que se forma durante el desarrollo embrionario. En el adulto este tejido se diversifica considerablemente, dando lugar a una gran variedad de tejidos diferentes. Una posible clasificación de los conectivos adultos es la que los diferencia según sus características mecánicas:
- Las fibras colágenas están formadas por colágeno, la proteína más abundante en el organismo. Son fuertes y resistentes a la tracción, pero flexibles, y se encuentran en la mayoría de los tejidos conectivos, especialmente en los más resistentes.
- Las fibras elásticas están formadas por dos tipos de proteínas, la elastina y la fibrilina. Pueden llegar a estirarse hasta el 150% de su longitud y luego recuperar su tamaño normal. Son más frecuentes en la piel, los vasos sanguíneos y el pulmón.
- Las fibras reticulares son haces finos de colágeno recubierto que soportan los vasos sanguíneos y otros tejidos conectivos. Soportan muchos órganos blandos y foman parte de las membranas basales.

- Tejido conectivo fibroso
- Conectivo laxo
- Tejido adiposo
- Conectivo denso
- Cartílago
- Tejido óseo
- Tejido conectivo líquido
- Sangre
- Linfa

El tejido conectivo denso presenta una gran cantidad de fibras colágenas agrupadas formando haces. Existen dos subtipos, el regular, que forma los tendones y los ligamentos, en el que los haces de fibras están alineados, y el irregular, que forma parte de la capa más profunda de la piel y en el que los haces no están organizados, sino que se distribuyen de modo aleatorio.

El tejido conectivo laxo se encuentra habitualmente entre otros tejidos, o entre órganos, manteniéndolos unidos entre sí. Sus células están separadas por una matriz gelatinosa rica en fibras colágenas y elásticas.

En el tejido adiposo los fibroblastos aumentan de tamaño y acumulan lípidos en su interior. En este caso la matriz extracelular es muy poco importante. Este tejido es un buen aislante térmico, de modo que se sitúa en la capa profunda de la piel para evitar la pérdida de calor. También constituye la principal reserva energética del organismo y participa en el dimorfismo sexual.
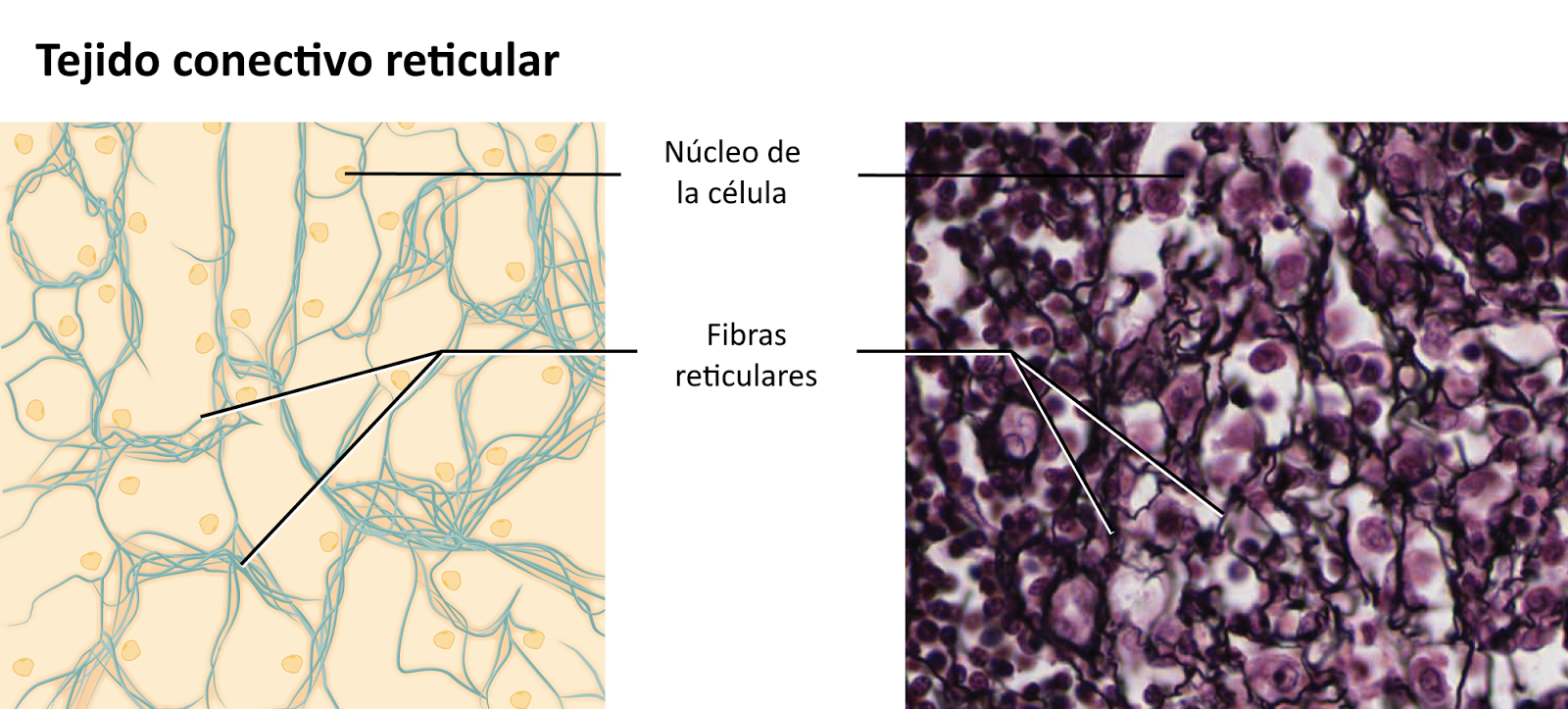
El tejido reticular posee únicamente ese tipo de fibras. Se denomina también tejido linfático porque es el principal constituyente de los órganos linfáticos (ganglios, timo, bazo y médula ósea roja) donde produce o almacena glóbulos blancos.
El tejido cartilaginoso se caracteriza al microscopio porque sus células se encuentran en cámaras llamadas lacunae (lagunas) y separadas entre sí por una matriz sólida pero flexible. Se pueden distinguir tres tipos de cartílago, según qué tipo de fibras sean las predominantes. El cartílago hialino presenta solo fibras de colágeno, y es el tipo más común, ya que se presenta en la nariz, la tráquea, las costillas, los extremos de los huesos, el esqueleto fetal... El fibrocartílago, por su parte, posee una gran cantidad aún mayor de fibras de colágeno, lo que le proporciona una gran resistencia. Se presenta en los discos intervertebrales o en la rodilla. Por último, el cartílago elástico posee también una cantidad considerable de fibras elásticas y se encuentra, por ejemplo, en la oreja.


El tejido óseo se organiza en los huesos de dos modos diferentes. El tejido óseo compacto ocupa la parte externa de los huesos. Está formado por unidades cilíndricas llamadas osteonas o sistemas de Havers que dejan en su centro un canal hueco, llamado conducto de Havers, por el que circulan nervios y los vasos sanguíneos que proporcionan nutrientes a las células. La matriz del tejido se sitúa alrededor de los conductos de Havers formando capas concéntricas llamadas lamelae (lamelas) entre las que se sitúan los osteocitos.


Se trata de un tejido menos macizo que el hueso compacto, a pesar de lo cual es capaz de soportar bien esfuerzos considerables gracias a su estructura semejante a las vigas que forman un edificio.

Existen dos tipos de tejido que pueden ocupar los espacios entre las trabéculas del hueso esponjoso. La médula ósea amarilla es básicamente un tejido adiposo con algunos eritrocitos dispersos en ella, mientras que la médula roja es un órgano hematopoyético, es decir, se encarga de producir las células de la sangre.

Membranas
En Anatomía se emplea este término para designar a láminas de tejido flexible que recubren alguna parte del cuerpo, mientras que el término epitelio (no tejido epitelial) se usa para denominar a la combinación de una capa de tejido epitelial y el tejido conectivo situado bajo ella. En el organismo podemos encontrar varios tipos de membranas:
- Las membranas mucosas tapizan cavidades que se abren al exterior del cuerpo, como las del digestivo, el respiratorio, el reproductor o el urinario. Las uniones estrechas entre las células epiteliales forman una barrera que es difícil de superar, lo que hace que estas membranas se comporten como buenos aislantes. Además segregan moco, que evita la deshidratación del tejido y ayudan a lubricar el epitelio. Puesto que, como todos los tejidos epiteliales, las mucosas carecen de vasos sanguíneos, se nutren a través de su membrana basal con nutrientes procedentes del conectivo subyacente.
- Las membranas serosas, por su parte, tapizan las cavidades corporales que no se abren al exterior y también recubren los órganos que se encuentran en esas cavidades. Están formadas por una capa de conectivo laxo sobre el que descansa un epitelio simple, que recibe el nombre de mesotelio. Los mesotelios segregan un líquido seroso, de composición semejante a la del suero sanguíneo, que lubrica estos tejidos facilitando el movimiento de los órganos. Las serosas que recubren las cavidades corporales tienen dos capas, la parietal y la visceral, entre las que hay una cierta cantidad de líquido.
- Las membranas sinoviales se encuentran revistiendo las cavidades articulares como la rodilla. Estas membranas no tienen tejido epitelial, sino que presentan, hacia el espacio articular, una capa discontinua de células llamadas sinoviocitos. Por debajo aparece un conectivo con adipocitos y fibras colágenas. El líquido sinovial lubrica y nutre al cartílago, y contiene macrófagos que eliminan microorganismos y restos celulares dañados.
- La piel es también una membrana, a la que se suele llamar membrana cutánea. Consta de un epitelio pluriestratificado y queratinizado, es decir, que acumula en sus capas externas, formadas por células muertas, una proteína llamada queratina. Por debajo del epitelio aparece un tejido llamado dermis, formado por conectivo areolar y conectivo denso irregular.
Es un tipo de tejido totalmente especializado en la producción de movimiento mediante el acortamiento de sus células. Dichas células presentan una morfología y una estructura interna bastante diferentes a las del resto del organismo, a pesar de que sus orgánulos sean exactamente los mismos, y reciben el nombre de fibras musculares.

Existen tres tipos de tejido muscular, que se diferencian entre sí por la forma de sus células, por su aspecto microscópico y por las características de su funcionamiento.

Las células que lo componen, llamadas fibras musculares esqueléticas, son pluricelulares, de forma cilíndrica y de gran longitud. Se forman durante el desarrollo embrionario mediante la fusión de varias células uninucleadas.
La contracción del músculo esquelético está siempre bajo control voluntario del sistema nervioso.

Las células del tejido muscular liso son diferentes a las que forman el músculo estriado. Son más pequeñas, unicelulares y con forma de huso. En ellas los elementos contráctiles no se disponen en sarcómeros, sino formando una red justo por debajo de la membrana plasmática, que constriñe a la célula en el momento de la contracción.
La contracción del músculo liso hace posible, por ejemplo, la circulación de la sangre en las arterias o el avance del alimento a lo largo del tubo digestivo, gracias a su actividad peristáltica.

Al microscopio se observan estriaciones similares a las del músculo esquelético, aunque sus células son diferentes a las de aquel, ya que son ramificadas y unicelulares, aunque están íntimamente unidas entre sí por medio de uniones especializadas que permiten la transmisión muy rápida del impulso contráctil a través de las propias fibras.
Tejido nervioso


En general las neuronas no llegan a estar conectadas físicamente, sino que en la mayoría de los casos queda entre ellas un pequeño espacio llamado espacio sináptico. La sinapsis es un tipo de unión especializada que permite la comunicación entre células cercanas por medio de la difusión de sustancias químicas llamadas neurotransmisores que son liberadas por el axón de la neurona presináptica y que activan receptores en la neurona postsináptica.
Células excitables

En las células musculares la llegada de un potencial de acción provoca la contracción de la célula debido a cambios de posición de las proteínas de su citoesqueleto (actina y miosina).
En las neuronas, la llegada de un neurotransmisor a una de sus dendritas o de un potencial de acción (existen también sinapsis eléctricas, en las que el contacto entre célula y célula se produce mediante uniones celulares que permiten el paso de iones) provoca el cambio en las propiedades de la membrana. La entrada de iones de sodio cambia la diferencia de carga que hay entre el interior y el exterior de la célula, proceso que se va "transmitiendo" a lo largo de la célula hasta llegar al extremo del axón. En ese punto el cambio de carga da lugar a la liberación de neurotransmisores que, cuando alcanzan la siguiente neurona, desencadenan o inhiben un nuevo potencial de acción en ella.
Reparación y regeneración tisular

En estos procesos pueden participar dos tipos de tejidos:
- El parénquima, es decir, el tejido activo del órgano. Cuando esto ocurre el nuevo tejido formado es plenamente funcional.
- El estroma, es decir, el tejido conectivo próximo al órgano. En este caso la parte que se regenera no tiene capacidad para realizar las funciones normales, por lo que se produce un tejido cicatricial.
Algunos tejidos conectivos conservan una gran capacidad de regeneración, como ocurre con el hueso, que se encuentra en un proceso de remodelación continuo llevado a cabo por los osteoblastos y los osteoclastos, que se encargan de eliminar la matriz en las zonas en las que no es necesaria. También la médula ósea es un tejido en regeneración permanente para dar lugar de forma continua a las células sanguíneas que van degradándose con el paso del tiempo. En este caso intervienen las células madre hematopoyéticas.
En cambio, otros tejidos conectivos como el cartílago se regeneran muy poco debido a su escasa irrigación, que dificulta la llegada de los nutrientes necesarios para el crecimiento celular.
Los tejidos musculares tienen, en general, una capacidad de regeneración muy baja. El músculo esquelético conserva células madre, denominadas células satélite, pero su división es demasiado lenta para poder reemplazar a las fibras dañadas. Las fibras musculares lisas son capaces de dividirse, pero lo hacen a un ritmo mucho más bajo que las células epiteliales o conectivas. Por su parte el músculo cardiaco carece de células madre y sus fibras son incapaces de dividirse. A pesar de ello se ha observado cierta capacidad de regeneración, que se debe a la llegada e infiltración en el tejido de células madre procedentes del sistema cardiovascular, que son capaces de diferenciarse en fibras musculares y células endoteliales.
Finalmente el tejido nervioso conserva algunas células madre, pero no ha sido posible encontrar pruebas de regeneración in vivo.